kanin. 4.21. Ang posisyon ng electrical axis ng puso ay pahalang (anggulo a * +15*). Mayroon ding pag-ikot ng puso sa paligid ng longitudinal axis na counterclockwise (complex QRS sa mga lead V 5 at V 6 na uri QR, transition zone (TZ) sa lead V 2 .
kanin. 4.22. Mayroong pag-ikot ng electrical axis ng puso sa kanan (anggulo a * +120°), pati na rin ang pag-ikot ng puso sa paligid ng longitudinal axis clockwise PZ sa lead V 6 (complex QRS sa mga lead V 5 (V 6 type AS).
Pagpapasiya ng pag-ikot ng puso sa paligid ng transverse axis (apex forward o backward)
Hindi gaanong karaniwan, ang ECG ay nagtatala ng mga pag-ikot ng puso sa paligid ng transverse axis nito, na nagaganap sa anteroposterior (sagittal) na eroplano (Larawan 4.23). Ang mga pag-ikot ng puso sa paligid ng transverse axis ay kadalasang nauugnay sa paglihis ng tuktok ng puso pasulong o paatras.
na may kaugnayan sa karaniwang posisyon nito, na humahantong sa isang paglabag sa karaniwang spatial na pag-aayos ng tatlong sandali na mga vectors ng ventricular depolarization sa sagittal at frontal na mga eroplano. Ang mga pag-ikot ng puso sa paligid ng transverse axis na may tuktok na pasulong o paatras ay pinakamahusay na naitala sa tatlong karaniwang limb lead. Tingnan ang fig. 4.23. Inilalarawan nito ang pamilyar na anim na axis na Bailey coordinate system, na pinaikot sa isang tiyak na anggulo sa tagamasid, pati na rin ang spatial na pag-aayos ng tatlong sandali ng mga vectors (0.02 s, 0.04 s at 0.06 s).
Sa karamihan ng mga kaso, na may normal na posisyon ng puso (Larawan 4.23, a), ang paunang torque vector (0.02 s) ay bahagyang naka-orient pataas at pakanan, at ang huling torque vector (0.06 s) ay naka-orient pataas at sa ang kaliwa o kanan. Ang parehong mga vector ay spatially na matatagpuan sa isang tiyak na anggulo sa pangharap na eroplano, na may 0.02 s vector na nakatuon sa pasulong, at ang 0.06 s na vector ay paatras. Ang parehong mga vector ay inaasahang papunta sa mga negatibong bahagi ng mga palakol ng karaniwang mga lead, bilang isang resulta kung saan ang medyo maliit na amplitude na Q at Q wave ay maaaring maitala sa mga lead na ito. S. Dapat tandaan na ang mga ngipin Q At S maitatala lamang sa isa o dalawa sa tatlong karaniwang lead: sa I at II o sa II at III.
tip muna(Larawan 4.23, b) ang inisyal na moment vector (0.02 s) ay lumilipat pa pataas at bahagyang pakanan, at samakatuwid ang ngipin Q nagsisimulang mairehistro sa lahat ng tatlong karaniwang lead at nagiging mas malinaw.
Ang huling sandali ng vector (0.06 s) ay lumilihis pababa at pabalik, bilang isang resulta kung saan ito ay matatagpuan halos patayo sa frontal na eroplano. Samakatuwid, ang projection nito sa mga axes ng lahat ng standard na lead ay lumalapit sa zero, na humahantong sa pagkawala ng wave 5 sa mga lead na ito.
Kapag umiikot ang puso sa paligid ng transverse axis tip pabalik(Larawan 4.23, c) ang inisyal na moment vector (0.02 s) ay lumilipat pasulong at pababa upang ang oryentasyon nito sa espasyo ay lumabas na halos patayo sa frontal plane. Samakatuwid, ang projection ng 0.02 s vector sa axis ng karaniwang mga lead ay lumalapit sa zero, at ang mga ngipin mismo Q ay hindi nakarehistro.
Ang panghuling sandali ng vector (0.06 s) ay lumilipat nang higit pa paitaas at nagsisimulang maipakita sa mga negatibong bahagi ng mga palakol ng lahat ng tatlong karaniwang limb lead, na humahantong sa hitsura ng medyo malalalim na ngipin. S v S u At Sm.
Kaya, upang matukoy ang pag-ikot ng puso sa paligid ng transverse axis, kinakailangan upang suriin ang pagsasaayos ng complex QRS sa karaniwang limb lead.
Pagsusuri ng Atrial Wave R
Matapos matukoy ang pag-ikot ng puso sa paligid ng anteroposterior, longitudinal at transverse axes, magpatuloy sa pagsusuri ng atrial wave R. Prong analysis R kabilang ang: 1) pagsukat ng amplitude ng ngipin R, 2) pagsukat ng tagal ng ngipin RU 3) pagpapasiya ng polarity ng ngipin R, 4) pagpapasiya ng hugis ng ngipin R.
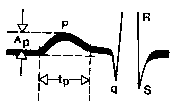
Prong amplitude R ay sinusukat mula sa contour line hanggang sa tuktok ng prong, at ang tagal nito ay mula sa simula hanggang sa dulo ng prong, tulad ng ipinapakita

sa Fig. 4.24. Normal na amplitude ng ngipin R ay hindi hihigit sa 2.5 mm, at ang tagal nito ay 0.1 s. Prong polarity R sa mga lead I, II at III ay ang pinakamahalagang electrocardiographic sign, na nagpapahiwatig ng direksyon ng paggalaw ng alon ng paggulo sa kahabaan ng atria at, dahil dito, ang lokalisasyon ng pinagmulan ng paggulo (pacemaker). Tulad ng naaalala mo, sa normal na paggalaw ng excitation wave sa kahabaan ng atria mula sa itaas hanggang sa ibaba at sa kaliwa, ang mga ngipin ay positibo, at kapag ang paggulo ay nakadirekta mula sa ibaba hanggang sa itaas, sila ay negatibo. Sa huling kaso, ang pacemaker ay matatagpuan sa ibabang bahagi ng atria o sa itaas na bahagi ng AV. node. Sa paggulo na nagmumula sa gitnang bahagi ng kanang atrium, ang depolarization wave ay nakadirekta sa parehong pataas at pababa. Average na vector R nakadirekta sa kaliwa, ayon sa pagkakabanggit, ang ngipin R ( lumalaki, nagiging mas malaki ang ngipin Plv at ang P wave ||(nagiging negatibo at mababaw.
Ang pagtukoy sa hugis ng ngipin ay may malaking kahalagahan R. Nahati sa dalawang tuktok, lumawak na ngipin R sa kaliwang mga lead (I, aVL, V 5, V 6) ay tipikal para sa mga pasyenteng may mga depekto sa mitral na puso at kaliwang atrial hypertrophy, at matulis na mataas na amplitude na ngipin R sa mga lead I, III, aVF ay sinusunod na may hypertrophy ng kanang atrium sa mga pasyente na may cor pulmonale (para sa higit pang mga detalye, tingnan ang Kabanata 7).
Pagsusuri ng kumplikadong ventricular QRST
Inflorescence ay isang shoot o sistema ng mga shoots na nagdadala ng mga bulaklak. Sa mga node ng inflorescence axes mayroong parehong mga dahon tulad ng sa vegetative na bahagi ng shoot, o binago ang mga nawalan ng kakayahang mag-photosynthesize - bracts, at sa mga node ng pedicel - bracts.
Ang biological na bentahe ng mga inflorescences sa mga solong bulaklak ay upang madagdagan ang garantiya ng polinasyon at upang mabawasan ang posibilidad ng pinsala sa mga bulaklak sa pamamagitan ng hindi kanais-nais na mga kadahilanan sa kapaligiran dahil sa kanilang unti-unting pamumulaklak. Karamihan sa mga halaman ay may mga inflorescence.Mayroong dalawang uri ng mga inflorescence: kumplikado, kapag ang mga bulaklak ay matatagpuan sa mga sanga ng pangunahing aksis, at simple lang, kapag ang mga bulaklak na mayroon o walang pedicels ay matatagpuan nang direkta sa pangunahing axis.
Ang mga kumplikadong inflorescence, depende sa paraan ng paglaki ng mga palakol, ay nahahati sa: sympodial(tinukoy) - ang axis ay nagtatapos sa isang bulaklak, ang mga bulaklak ay namumulaklak mula sa itaas hanggang sa mga sanga sa gilid o sentripugal kung ang mga bulaklak ay matatagpuan sa parehong eroplano; At monopodial(indefinite) - ang axis ay lumalaki nang walang katiyakan, ang mga bulaklak ay namumulaklak mula sa base hanggang sa tuktok o centripetally kung ang mga bulaklak ay matatagpuan sa parehong eroplano.
Mga kumplikadong sympodial inflorescences:
- monochasium - ang pangunahing axis ay nagtatapos sa isang bulaklak; isang axis ng pangalawang pagkakasunud-sunod ay nabuo sa ilalim nito, nakoronahan din ng isang bulaklak, atbp.; kung ang mga subfloral axes ay gumagalaw sa isang direksyon, pagkatapos ay isang curl ay nabuo, ngunit kung halili sa isang direksyon o sa iba pa, isang gyrus ay nabuo; isang kulot kung saan pinaikli ang mga lateral branch ay tinatawag na glomerulus;
- dichasia - dalawang magkasalungat na palakol ay nabuo sa ilalim ng bulaklak, ang bawat isa ay nagtatapos din sa isang bulaklak at gumagawa din ng dalawang subfloral axes, paulit-ulit ang parehong paraan ng pagsasanga, atbp.;
- pleiochasia - higit sa dalawang subfloral axes ay umaabot mula sa pangunahing axis, na nagdadala ng isang apical na bulaklak, na bumubuo ng isang whorl ng monochasia o dichasia;
- thyrsus - ang mga inflorescences sa itaas ay matatagpuan sa pangunahing axis, kadalasan ay may hugis na pyramidal.
Mga kumplikadong monopodial inflorescences:
- panicle - isang napaka branched inflorescence, ang mas mababang lateral na mga sanga ay mas malakas na sangay kaysa sa itaas;
- corymb - isang panicle kung saan ang mga bulaklak ay matatagpuan higit pa o mas kaunti sa parehong eroplano;
- kumplikadong spike - ang pangunahing axis ay bumubuo ng mga sanga kung saan matatagpuan ang mga bulaklak na walang pedicels; ang mga sanga na ito ay tinatawag na mga spikelet;
- kumplikadong payong - ang mga distansya sa pagitan ng mga second-order axes ay pinaikli, at lumayo sila mula sa tuktok ng first-order axis; ang mga distansya sa pagitan ng mga pedicels ay pinaikli, at sila ay matatagpuan sa tuktok ng pangalawang-order na axis; Kadalasan ang mga dahon sa base ng second-order axes ay bumubuo ng isang pangkalahatang involucre, at sa base ng pedicels - isang pribadong involucre.
Bilang karagdagan sa mga nakalista, may mga kumplikadong inflorescence na tinatawag pinagsama-sama. Ang mga ito ay nabuo sa pamamagitan ng isang kumbinasyon ng iba't ibang uri ng mga inflorescence. Halimbawa, ang yarrow ay may pinagsama-samang inflorescence - mga basket na nakolekta sa isang kalasag; Ang fescue at bluegrass ay may mga spikelet na nakolekta sa isang panicle.
Mga simpleng inflorescences na may pinahabang axis:
- raceme - ang pangunahing axis ay nagdadala ng mga bulaklak na may mga pedicels, kadalasang may parehong haba;
- spike - ang pangunahing axis ay nagdadala ng mga bulaklak na walang pedicels;
- catkin - isang nakabitin na tainga, iyon ay, isang tainga na may malambot na axis; pagkatapos ng pamumulaklak ang inflorescence ay karaniwang nahuhulog;
- cob - isang tainga na may malakas na makapal na axis, na napapalibutan ng isa o higit pang mga dahon, ang tinatawag na spathe, o pakpak.
Mga simpleng inflorescences na may pinaikling axis:
- payong - isang inflorescence kung saan ang pangunahing axis ay pinaikli, at ang mga pedicels, na may halos parehong haba, ay umaabot mula sa tuktok ng axis;
- ulo - isang payong na ang mga bulaklak ay alinman sa walang pedicels o napakaikli;
- basket - ang tuktok ng pangunahing axis ay lumalaki sa anyo ng isang kama kung saan matatagpuan ang mahigpit na saradong mga bulaklak; ang mga apikal na dahon ay masikip at bumubuo ng isang involucre.
Mga inflorescences ng cymoid ay isang malaking grupo ng mga inflorescence. Kabilang sa mga cymoid inflorescences, mayroong dalawang pangunahing uri: cymoids at thyrsi. Ang mga cymoid ay karaniwang pinasimple na thyrsi. Ang Thyrsae ay mga branched inflorescences, at ang antas ng pagsanga ay bumababa mula sa base hanggang sa tuktok. Ang pangunahing axis ng thyrsus ay lumalaki nang monopodially, ngunit ang mga bahagyang inflorescences ng isang order o iba pa ay cymoids.
Ang Thyrsae na ang pangunahing axis ay nagtatapos sa isang bulaklak ay tinatawag na sarado, kung hindi, sila ay itinuturing na bukas. Depende sa antas ng pagsasanga ng mga lateral axes, ang pleiothyrsus ay nakikilala, kung saan ang mga cymoid ay matatagpuan sa mga axes ng ikatlo at mas mataas na mga order; dithyrs, kung saan matatagpuan ang mga cymoid sa mga palakol ng pangalawang pagkakasunud-sunod; at monothyrsus, kung saan matatagpuan ang mga cymoid nang direkta sa pangunahing axis ng inflorescence. hugis payong, capitate thyrsi, atbp. Ang Thyrsae ay madaling ma-convert sa iba pang mga uri ng mga inflorescences sa pamamagitan ng pagbawas (pagbaba) ng bilang ng mga lateral axes, pagpapaikli ng internodes, atbp. Ang thyrsae ay madalas na matatagpuan sa mga halaman. Halimbawa, ang thyrsus ay ang inflorescence ng horse chestnut. Ang mga thyrs ng iba't ibang uri ay kumakatawan sa mga inflorescences ng maraming Lamiaceae. Ang mga inflorescences ng Birch ay hugis-catkin na thyrsus.
Ang subfamily ng sedges proper (Caricoideae) ay nailalarawan sa pamamagitan ng unisexual na mga bulaklak na walang perianth, maliban sa Siberian cobresia (Kobresia sibirica), kung saan ito ay binubuo ng tatlong brown na kaliskis (Fig. 167). Ang mga spikelet ng mga kinatawan ng pamilyang ito ay nakaupo sa mga axils ng scaly covering dahon at napapalibutan ng mga kaliskis na matatagpuan sa ventral (ventral) na bahagi ng spikelet at homologous sa preleaf ng vegetative shoot. Sa genera na Schoenoxiphium at Kobresia, ang mga kaliskis na ito ay may mga unfused edge o hanggang kalahati (bihirang sa tuktok) fused edge, at sa genera na Uncinia at Carex, ang mga ito ay lubos na metamorphosed, sarado, at sac-shaped. Ang Schenoxyphium at cobresia ay karaniwang may mga bisexual spikelet na may pinahaba (sa unang genus) at isang napakaikli, halos hindi nabuo (sa pangalawang genus) axis. Sa uncinia at sedge, ang mga spikelet ay nabawasan sa isang babaeng bulaklak, na nakapaloob sa isang parang pouch na sukat na tinatawag na pouch. Sa mga species ng Uncinia, ang babaeng bulaklak ay nakaupo sa base ng napanatili pa ring spikelet; ang huli ay ganap na nabawasan sa karamihan ng mga sedge. Stamens 3. Estilo na may 2 - 3 stigmatic na sanga. Ang subfamily ay binubuo ng isang sedge tribe (Cariceae), kabilang ang 4 na genera. Ang pinaka-primitive sa kanila ay Schenoxyphium, na may bilang na mga 15 species, kung saan 2 ay lumalaki sa Madagascar, at ang natitira sa South Africa. Lumalaki sila sa mga bundok, sa mamasa-masa at latian na mga lugar. Ito ay mga halamang pangmatagalan na may basal, kadalasang mga patag na dahon at hugis spike o paniculate inflorescence. Ang mga spikelet ay napapalibutan ng iba't ibang antas ng saradong, minsan parang saccule na kaliskis. Sa loob nito ay may isang malakas na flattened axis ng spikelet, sa base kung saan nakaupo ang isang babaeng bulaklak, at sa tuktok mayroong ilang mga lalaki (Larawan 167). Sa ilang mga species ang axis ay hindi nagdadala ng mga lalaki na bulaklak; ang huli ay bumubuo ng mga lalaking spikelet sa parehong inflorescence.
Ang susunod na genus ng sedge subfamily - Uncinia - ay binubuo ng 40 - 50 species, na ibinahagi pangunahin sa southern hemisphere (maliban sa Africa). Karamihan sa mga species (32) ay katutubong sa New Zealand. Hilaga ng ekwador, ang uncinia ay matatagpuan sa Timog-silangang Asya (Kalimantan Island, Pilipinas), Hawaiian Islands, Central at South America at West Indies. Ang mga species ng genus na ito ay lumalaki mula sa antas ng dagat hanggang sa alpine belt, sa marshy meadows, swamps, mamasa kagubatan at bushes, kung saan madalas silang bumubuo ng mga thicket. Ang mga species ng Uncinia ay pangmatagalan, kadalasang siksik na turfy herbs na may basal linear na dahon. Ang inflorescence ay isang apikal na spike na nagtataglay ng mga lalaking bulaklak sa itaas na bahagi, at mga sac na nakapaloob sa mga babaeng bulaklak sa ibabang bahagi, kung saan nakausli ang isang hugis kawit na spikelet axis na nakakurbada sa tuktok. Ang hugis-kawit na tuktok ng axis ay isang matibay, nakabalot na pantakip na sukat, sa sinus kung saan, tulad ng ipinakita ng mga anatomical na pag-aaral, mayroong isang hindi maunlad na bato. Stamens 3. Estilo na may 3 sanga na hugis stigma. Ang prutas ay tatsulok, nakapaloob sa isang supot. Sa kanilang hugis-kawit na axis, ang mga sako ay kumakapit sa balahibo o balahibo ng mga hayop at sa gayon ay kumalat.
Kasama sa genus Cobresia ang humigit-kumulang 40 species, na pangunahing ipinamamahagi sa mga kabundukan ng mapagtimpi at bahagyang subtropiko at tropikal na mga sona ng hilagang hemisphere, ngunit pangunahin sa Asya; ilang species ang tumagos sa Arctic. Ang Cobresias ay mga katangiang halaman ng kabundukan na may mababang pag-ulan. Lumalaki sila sa tabi ng mga pampang ng mga ilog at lawa, mamasa-masa at latian, kadalasang maalat na parang, parang-steppe at mabatong mga dalisdis. Ang mga species ng genus na ito ay kadalasang bumubuo ng cobresia meadows (tinatawag na cobresia meadows o heathlands). Ang Cobresias ay mga perennial dense-turf grass na may mga basal na dahon na may flat o brush-like plates. Ang inflorescence ay hugis spike o mas madalas na paniculate, ay binubuo ng maliit na 2-7-flowered bisexual spikelets, ang pinakaibabaw nito ay kung minsan ay lalaki. Ang mga kaliskis na nakapalibot sa spikelet ay kadalasang may hindi magkadikit na mga gilid o, bihirang, pinagsamang mga gilid hanggang kalahati o halos sa itaas. Ang mga spikelet (ang kanilang axis ay lubos na pinaikli, halos hindi ipinahayag) ay binubuo ng isang babaeng bulaklak at 1 - 6 na lalaki na bulaklak na matatagpuan sa labas mula dito. Ang huli ay binubuo ng 3 stamens na nakaupo sa axil ng pantakip na kaliskis. Istilo na may 3, bihirang 2 stigmatic na sanga. Karaniwang tatsulok ang prutas. Ang mga Cobresia ay nagpaparami sa pamamagitan ng mga buto, at isa lamang sa kanila - malakas na cobresia (K. robusta) - din vegetatively, sa pamamagitan ng gumagapang na rhizomes. Ang mga prutas ay ipinamamahagi ng mga herbivorous mammal. Cobresia mabalahibo, (K. capilliformis), na sumasakop sa malawak na mga lugar sa Tien Shan at Pamir-Alai, pati na rin ang isang bilang ng iba pang mga species ay mahalagang pastulan halaman.

Ang genus sedge (Carex) ay may humigit-kumulang 1500 (ayon sa iba pang pinagkukunan hanggang 2000 - 2500) species at nabibilang sa kakaunting pinakamalaking genera ng angiosperms. Ang mga species ng sedge ay laganap sa buong mundo, ngunit higit sa lahat sa mapagtimpi at malamig na mga rehiyon, kung saan gumaganap sila ng mahalagang papel sa komposisyon ng vegetation cover ng hindi lamang mamasa-masa at latian, kundi pati na rin ang mga tuyong tirahan. Sa mga tuntunin ng antas ng pakikilahok sa vegetation cover ng Unyong Sobyet, kung saan matatagpuan ang humigit-kumulang 400 species ng mga sedge, sila ay pangalawa lamang sa mga damo at asteraceae. Maraming mga species ng genus na ito ang mga tagapagturo ng iba't ibang mga komunidad ng halaman, kabilang ang steppe at kahit semi-disyerto at disyerto. Ang mga sedge ay lumalaki halos saanman, na nagaganap mula sa antas ng dagat hanggang sa matataas na bundok. Karamihan sa mga species ay naninirahan sa mga latian, latian at mamasa-masa na parang, at sa mga pampang ng mga anyong tubig (halimbawa, bote sedge - C. rostrata, bladder sedge - C. vesicaria, fox sedge - C. vulpina, Fig. 174, dioecious sedge - C. dioica, Fig. 174). Ang iba pang mga sedge ay matatagpuan sa iba't ibang uri ng kagubatan, kabilang ang mga tropikal na maulang kagubatan. Ang ilang mga species ay lumalaki sa mga tuyong parang, steppes, at tuyong mga dalisdis (mababang sedge - C. hnmilis, maagang sedge - C. paecox, squat sedge - C. supina, atbp.). Ang makapal na columnar sedge (C. pachystylis) ay isang katangian ng halaman ng clayey semi-deserts, at ang namamaga na sedge (C. physodes) ay isang katangian ng halaman ng mabuhangin na disyerto. Ang huling species ay may kakaibang orihinal na anyo dahil sa malakas na parang bula, mapula-kalawang, napakalaki (hanggang 2 cm ang haba) na mga sako na namamaga ng prutas (Fig. 174). Ilang uri ng hayop ang naninirahan sa mga baybayin ng dagat, buhangin, mga parang sa baybayin, maliliit na bato, at mga bato. Karamihan sa mga species sa temperate zone ay lumalaki sa kapatagan, ngunit ang ilang mga sedge ay karaniwang mga naninirahan sa subalpine at alpine meadows. Sa mga tropiko, lalo na sa rehiyon ng floristic ng Malesian, kung saan lumalaki ang 66 na species ng mga sedge, ang huli ay naninirahan pangunahin sa mga bundok at mga 10 species lamang ang lumalaki mula sa antas ng dagat hanggang 500 m sa itaas ng antas ng dagat.
Ang mga sedge ay pangmatagalan na monoecious, napakabihirang dioecious, mga halamang gamot na may gumagapang o pinaikling rhizome, na bumubuo ng mga tuft o kung minsan ay hummocks. Ang mga dahon ay karaniwang lahat ay basal, mas madalas na mayroon ding mga dahon ng stem na umaabot mula sa mga node na matatagpuan halos kasama ang buong stem; Ang talim ng dahon ay karaniwang linear, bihirang lanceolate (halimbawa, sa kalawang-batik-batik na sedge - C. siderosticta, Fig. 175), at kung minsan ay makitid sa isang tangkay. Ang mga bulaklak ay unisexual, sa mga spikelet na bumubuo ng hugis spike, paniculate, racemose o capitate inflorescence; minsan ang inflorescence ay single-spikelet. Ang mga spikelet ay ganap na binubuo ng lalaki o babaeng bulaklak o pinaghalo, ng dalawang uri: androgynous - na may mga lalaking bulaklak sa itaas na bahagi ng spikelet at babaeng bulaklak sa ibabang bahagi, gynecandrous - na may mga babaeng bulaklak sa tuktok, na may mga lalaki na bulaklak sa ang ilalim. Ang mga babaeng bulaklak ay nakapaloob sa isang sac na tumataas na may prutas, na nagpapaliit sa tuktok sa isang spout, mula sa kung saan ang bahagi ng estilo at stigmatic na mga sanga ay nakausli. Nasabi na sa itaas na ang mga babaeng bulaklak ng mga sedge, na nakapaloob sa mga sac, ay nabawasan, mga single-flowered spikelet. Samakatuwid, ang isang koleksyon ng mga sac na nakaupo sa parehong axis ay isang kumplikadong spike. Ang mga koleksyon ng mga lalaking bulaklak ay mga simpleng spike. Gayunpaman, para sa kaginhawahan, ang parehong kumplikado at simpleng mga spike ng mga sedge ay karaniwang tinatawag na mga spikelet. Ang mga character ng mga sac ay lubhang magkakaibang at may malaking kahalagahan para sa taxonomy ng mga sedge. Ang maraming mga species ng genus na ito ay naiiba pangunahin sa kanilang mga supot. Ang mga sac ay isang organ na walang alinlangan na may mahalagang biological na kahalagahan. Marahil ito ay salamat sa paglitaw ng sac na ang genus sedge ay naging pinakamarami at laganap sa lahat ng genera ng pamilya. Pinoprotektahan ng mga sac ang obaryo at ang nabubuong prutas mula sa pagkakalantad sa hindi kanais-nais na mga kondisyon, at sa maraming mga species sila ay isang adaptasyon din sa pamamahagi ng mga prutas. Ang estilo ng mga sedge ay may 2 - 3 karaniwang mahahabang sanga ng stigmatic. Ang mga prutas ay tatsulok o bahagyang biconvex.
Maraming sedge ang namumulaklak sa unang bahagi ng tagsibol. Ang mga species ng genus na ito ay mga anemophilous na halaman, ngunit ang ilan sa kanila ay tila nagpapakita rin ng entomophily. Kaya, ito ay kilala na ang dalawa sa mga pinakaunang namumulaklak na species - heath sedge (C. ericetorum) at carnation sedge (C. caryophyllea) - ay regular na binibisita ng honey bees upang mangolekta ng pollen, na tila nagsasagawa din ng cross-pollination. Halos walang impormasyon tungkol sa biology ng sedge flowering. Ang kanilang mga bulaklak ay itinuturing na protogynous. Ngunit, tulad ng ipinakita ng mga espesyal na obserbasyon ng pamumulaklak ng mabalahibong sedge (C. pilosa), ang morphological protogyny ay maaaring hindi tumutugma sa physiological (L. Antonova, 1976). Sa species na ito, ang hitsura ng ganap na binuo stigmas ay nauuna sa pagbubukas ng anthers. Gayunpaman, ang stigma ay umabot sa physiological maturity, ibig sabihin, ang kakayahang makita ang pollen, sa oras lamang na bumukas ang anthers at ang pollen ay nagkalat. Kaya, ang mabalahibong sedge ay mahalagang hindi isang prototypical, ngunit isang homogamous na halaman. Ang self-pollination ay malamang na hindi kasama ng bahagyang hindi pagkakatugma sa sarili. Ito ay eksperimento na itinatag na sa self-pollination ang species na ito ay gumagawa ng makabuluhang mas kaunting mga prutas kaysa sa cross-pollination. Ang mga sedge ay nagpaparami sa pamamagitan ng mga buto, ngunit sa mga species na may gumagapang na rhizomes, ang vegetative propagation ay kapansin-pansing nangingibabaw. Ang pamamahagi ng mga prutas ay nangyayari sa iba't ibang paraan. Ang mga bunga ng mga sedge sa baybayin, na nakapaloob sa mga sako na namamaga o espongha sa base, ay ipinamamahagi sa pamamagitan ng tubig. Ang mga sac ng namamagang sedge ay mahusay na iniangkop sa anemochory. Madali silang gumulong sa ibabaw ng buhangin kahit na sa mahinang hangin. Sa maraming mga species, ang mga sac ay ipinamamahagi nang epizoochorously - pangunahin na may dumi sa mga paa ng mga ibon. Ang mga sako ng ilang sedge ay maaari ding kumapit sa mga balahibo ng mga ibon. Ang endozoochorous na pamamahagi ng mga prutas kapag sila ay kinain ng mga itik ay may mahalagang papel. Ang mga ibon ay nagdadala ng orange-red, medyo may laman na mga sac ng tropikal na species ng sedge berry (C. baccans, Fig. 175). Ang mga sac ng ilang sedge sa kagubatan, tulad ng palmate sedge (C. digitata) at ornithopoda sedge (C. ornithopoda), na may mga pahabang laman na base, ay dinadala ng mga langgam.
Kapag umikot ang puso tugatog pasulong sa paligid ng transverse axis nito, ang average na QRS vector ay lumilihis pasulong, ang paunang vector (Q) ay nakadirekta nang higit pa sa kanan at pataas kaysa karaniwan (sa F plane). Ito ay matatagpuan parallel sa frontal plane at samakatuwid ay malinaw na mga proyekto sa minus axes ng lahat ng karaniwang mga lead (I, II at III).
Naka-on ECG isang binibigkas na alon QI, II, III ay naitala. Ang panghuling vector (S) ay lumilihis sa likuran at pababa, patayo sa pangharap na eroplano at hindi inaasahang minus sa axis ng karaniwang mga lead, samakatuwid, ang S wave ay hindi naitala sa mga lead I, II, III. Kaya, kapag ang puso umiikot sa tuktok nito pasulong sa paligid ng transverse axis sa ECG sa mga lead na I, II at III na nagtatala ng qR complex.
Kapag umikot ang puso kasama ang tuktok pabalik sa paligid ng transverse axis, ang average na QRS vector ay lumilihis pabalik (sa S plane), ang huling vector (S) ay lumilihis sa kanan at pataas, na nagbibigay ng isang makabuluhang projection sa negatibong poste ng mga axes ng lead I, II at III. Ang ECG ay nagpapakita ng isang binibigkas na alon SI, II, III. Ang paunang vector (Q) ay nakadirekta pababa at pasulong at samakatuwid ay hindi naka-proyekto sa negatibong poste ng mga axes ng karaniwang mga lead. Bilang resulta, walang Q wave sa ECG sa mga lead I, II at III. Ang QRSI, II, III complex ay kinakatawan ng uri ng RS.
ECG malusog na babae D., 30 taong gulang. Regular ang sinus ritmo, 67 kada minuto. P - Q=0.12 seg. P = 0.10 seg. QRS = 0.08 seg. Q - T = 0.38 seg. Ru>RI>Rir AQRS=+52°. Ap=+35°. Sa=+38°. Complex QRSI,II,III type qR. Ipinapakita nito na ang paunang vector (Q) ay nakadirekta sa kanan at pataas nang higit kaysa karaniwan, at samakatuwid ay inaasahang sa minus ng lahat ng karaniwang mga lead (wave qI, II, III). Ang pangwakas na vector (S) ay lumilihis sa likuran at pababa, patayo sa pangharap na eroplano at hindi nakaharap sa mga palakol ng mga lead I, II, III (walang S wave, cw). Ang ganitong mga pagbabago sa direksyon ng mga inisyal at panghuling vector ay maaaring dahil sa pag-ikot ng puso na may tuktok na pasulong. Dapat tandaan na ang QRS transition zone ay tumutugma sa lead V2, na siyang kanang hangganan ng normal na lokasyon nito. Complex QRSV5V6 type RS, na sumasalamin sa sabay-sabay na bahagyang pag-ikot clockwise sa paligid ng longitudinal axis. Ang P, T wave at ang RS - T segment ay normal sa lahat ng lead.
Konklusyon. Isang variant ng isang normal na ECG (pag-ikot ng puso na may tuktok na pasulong sa paligid ng transverse axis at clockwise sa paligid ng longitudinal axis).
ECG ng isang malusog na tao K., 37 taon. Malubhang sinus bradycardia, 50 bawat 1 min. Interval P - Q=0.15 seg. P = 0.11 seg. =0.09 seg. Q - T=0.39 seg. RII>RI>RIII. AQRS = +50°. Ar=+65°. Sa=+50°. Anggulo ng QRS - T=0°. Complex QRSI,II,III type qR. Ang Q wave ay pinaka-binibigkas sa lead II, kung saan ang amplitude nito ay 3 mm at ang tagal nito ay bahagyang mas mababa sa 0.03 sec. (normal na sukat). Ang inilarawang hugis ng QRS ay nauugnay sa pag-ikot ng puso sa tuktok nito pasulong.
Sa dibdib nangunguna ang QRSV5, V6 complex ay nasa uri din ng qR, at ang RV1 wave ay binibigkas, ngunit hindi pinalaki (amplitude 5 mm). Ang mga pagbabagong ito ng QRS ay nagpapahiwatig ng isang counterclockwise na pag-ikot ng puso sa paligid ng longitudinal axis nito. Ang transition zone ay karaniwang matatagpuan (sa pagitan ng V2 at V3). Ang natitirang mga alon ng ECG ay normal. Ang segment ng RS - TII,III ay nakataas ng hindi hihigit sa 0.5 mm, na maaaring normal.
Konklusyon. Sipon. Pag-ikot ng puso sa pakaliwa at sa tuktok na pasulong (isang variant ng isang normal na ECG).
ECG ng isang malusog na babae K., 31 taong gulang. Regular ang sinus ritmo, 67 kada minuto. P - Q=0.16 seg. P=0.09 seg. QRS=0.08 seg. Q - T=0.39 seg. RII>RI>RIII. AQRS=+56°. Sa=+26°. Anggulo ng QRS - T=30°. Ar=+35°.
Uri ng kumplikadong QRSI,II,III Rs. Ang binibigkas na S sa mga lead I, II, III ay nagpapahiwatig ng isang makabuluhang paglihis ng huling vector (S) sa kanan at pataas. Ang kawalan ng QI, II, III wave ay nauugnay sa direksyon ng paunang QRS vector pababa at pasulong (patungo sa positibong poste ng karaniwang mga lead). Ang oryentasyong ito ng mga inisyal at panghuling QRS vector ay maaaring dahil sa pag-ikot ng puso na ang tuktok nito ay paatras sa paligid ng transverse axis nito (type SI, SII, SIII ECG). Ang natitirang mga ECG wave ay nasa loob ng karaniwang normal na katangian: QRSV6 type qRs. Ang QRS transition zone sa pagitan ng V2 at V3, ang RS segment - TV2 ay inilipat paitaas ng 1 mm. Sa natitirang mga lead, ang RS-T ay nasa antas ng isoelectric na linya, ang TIII ay bahagyang negatibo, ang TaVF ay positibo, ang TV1 ay negatibo, ang TVJ_V6 ay positibo, na may bahagyang mas malaking amplitude sa V2V3. Ang P wave ay may normal na hugis at sukat.
Konklusyon. Variant ng normal na uri ng ECG SI, SII, SIII (pag-ikot ng puso na ang tuktok ay paatras sa paligid ng transverse axis).

Hindi gaanong karaniwan, ang ECG ay nagtatala ng mga pag-ikot ng puso sa paligid ng transverse axis nito, na nagaganap sa anteroposterior (sagittal) na eroplano (Larawan 57). Ang mga pag-ikot ng puso sa paligid ng transverse axis ay kadalasang nauugnay sa paglihis ng tuktok ng puso pasulong o paatras na may kaugnayan sa karaniwang posisyon nito, na humahantong sa isang paglabag sa karaniwang spatial na pag-aayos ng tatlong sandali na vectors ng ventricular depolarization sa sagittal at mga pangharap na eroplano.
Ang mga pag-ikot ng puso sa paligid ng transverse axis na may tuktok na pasulong o paatras ay pinakamahusay na naitala sa tatlong karaniwang limb lead. Tingnan ang fig. 57. Ipinapakita nito ang pamilyar na six-axis Bailey coordinate system, na pinaikot sa isang tiyak na anggulo sa tagamasid, pati na rin ang spatial na pag-aayos ng tatlong sandali ng mga vectors (0.02 s, 0.04 at 0.06 s).
SA
kanin. 57. ECG form sa tatlong karaniwang mga lead. a – sa normal na posisyon ng puso, b – kapag ang puso ay iniikot sa paligid ng transverse axis na may tuktok na pasulong; c – kapag ang puso ay iniikot sa paligid ng transverse axis na ang tuktok ay paatras.
Ang parehong mga vector ay spatially na matatagpuan sa isang tiyak na anggulo sa pangharap na eroplano, na may 0.02 s vector na nakatuon sa pasulong, at ang 0.06 s na vector ay paatras. Ang parehong mga vector ay naka-project sa mga negatibong bahagi ng mga axes ng karaniwang mga lead, bilang isang resulta kung saan ang medyo maliit na amplitude na Q at S wave ay maaaring maitala sa mga lead na ito. Dapat tandaan na ang Q at S waves ay maaaring maitala lamang sa isang o dalawa sa tatlong karaniwang lead: sa I at II o sa II at III.
Kapag ang puso ay umiikot sa paligid ng transverse axis na may tuktok na pasulong (Larawan 57, b), ang paunang torque vector (0.02 s) ay lumilipat nang mas paitaas at medyo pakanan, at samakatuwid ang Q wave ay nagsisimulang maitala sa lahat ng tatlo. karaniwang mga lead at nagiging mas malinaw.
Ang huling sandali ng vector (0.06 s) ay lumilihis pababa at pabalik, bilang isang resulta kung saan ito ay matatagpuan halos patayo sa frontal na eroplano. Samakatuwid, ang projection nito sa mga axes ng lahat ng standard na lead ay lumalapit sa zero, na humahantong sa paglaho ng S wave sa mga lead na ito.
TANDAAN! Kapag ang puso ay umiikot sa paligid ng transverse axis na may tuktok na pasulong, ang ventricular QRS complex sa mga karaniwang lead ay nasa anyo na qR I, qR II at qR III
Kapag ang puso ay umiikot sa paligid ng transverse axis na may tuktok na paatras (Larawan 57, c), ang inisyal na moment vector (0.02 s) ay lumilipat pasulong at pababa upang ang oryentasyon nito sa espasyo ay halos patayo sa frontal plane. Samakatuwid, ang projection ng 0.02 s vector sa axis ng karaniwang mga lead ay lumalapit sa zero, at ang Q waves mismo ay hindi naitala.
Ang huling sandali ng vector (0.06 s) ay lumilipat nang higit pa paitaas at nagsisimulang i-project sa mga negatibong bahagi ng mga palakol ng lahat ng tatlong karaniwang limb lead, na humahantong sa hitsura ng medyo malalalim na ngipin S I, S II at S III.
TANDAAN! Kapag ang puso ay umiikot sa paligid ng transverse axis na ang tuktok ay paatras, ang ventricular complex sa karaniwang mga lead ay nasa anyong RS I, RS II, RS III.
Kaya, upang matukoy ang pag-ikot ng puso sa paligid ng transverse axis, kinakailangan upang suriin ang pagsasaayos ng QRS complex sa karaniwang mga limb lead.
kanin. 58. Pagsukat ng amplitude at tagal ng P wave ng ECG. A p - amplitude ng P wave, t p - tagal ng P wave.