Pirinç. 4.21. Kalbin elektrik ekseninin konumu yataydır (a açısı * +15*). Ayrıca kalbin uzunlamasına eksen etrafında saat yönünün tersine bir dönüşü de vardır (karmaşık QRS V 5 ve V 6 tiplerinde derivasyonlarda QR, V 2'deki geçiş bölgesi (TZ).
Pirinç. 4.22. Kalbin elektrik ekseninin sağa doğru bir dönüşü vardır (a açısı * +120°) ve ayrıca V6 derivasyonunda (karmaşık) kalbin uzunlamasına eksen etrafında saat yönünde PZ dönüşü vardır. QRS V 5'te (V 6 tip AS).
Kalbin enine eksen etrafındaki rotasyonunun belirlenmesi (apeks ileri veya geri)
Daha az yaygın olarak, EKG, kalbin ön-arka (sagital) düzlemde meydana gelen enine ekseni etrafındaki rotasyonlarını kaydeder (Şekil 4.23). Kalbin enine eksen etrafındaki rotasyonları genellikle kalbin tepe noktasının ileri veya geri sapması ile ilişkilidir.
normal konumuna göre, bu da sagittal ve frontal düzlemlerde ventriküler depolarizasyonun üç moment vektörlerinin olağan mekansal düzenlemesinin ihlaline yol açar. Kalbin apeks ileri veya geri olacak şekilde transvers eksen etrafındaki rotasyonları en iyi üç standart ekstremite derivasyonunda kaydedilir. Şek. 4.23. Gözlemciye belirli bir açıyla döndürülmüş tanıdık altı eksenli Bailey koordinat sisteminin yanı sıra üç moment vektörünün (0,02 s, 0,04 s ve 0,06 s) uzamsal düzenlemesini gösterir.
Çoğu durumda, kalbin normal pozisyonunda (Şekil 4.23, a), ilk tork vektörü (0,02 s) hafifçe yukarı ve sağa doğru yönlendirilir ve son tork vektörü (0,06 s) yukarı ve sağa doğru yönlendirilir. sol veya sağ. Her iki vektör de uzaysal olarak ön düzleme belirli bir açıda yerleştirilmiştir; 0,02 s'lik vektör ileri doğru yönlendirilir ve 0,06 s'lik vektör geriye doğru yönlendirilir. Her iki vektör de standart uçların eksenlerinin negatif kısımlarına yansıtılır, bunun sonucunda bu uçlarda nispeten küçük genlikli Q ve Q dalgaları kaydedilebilir. S. Unutulmamalıdır ki dişler Q Ve Süç standart derivasyondan yalnızca bir veya ikisine kaydedilebilir: I ve II veya II ve III'te.
önce ipucu(Şekil 4.23, b) başlangıç moment vektörü (0,02 s) daha da yukarıya ve hafifçe sağa doğru kayar ve dolayısıyla diş Q her üç standart ipucunda da kaydedilmeye başlar ve daha belirgin hale gelir.
Son moment vektörü (0,06 s) aşağı ve geriye doğru sapar, bunun sonucunda artık ön düzleme neredeyse dik olarak konumlandırılır. Bu nedenle, tüm standart uçların eksenleri üzerindeki projeksiyonu sıfıra yaklaşır ve bu da bu uçlarda 5. dalganın kaybolmasına yol açar.
Kalp enine eksen etrafında döndüğünde geri bahşiş(Şekil 4.23, c) başlangıç moment vektörü (0,02 s) ileri ve aşağı doğru kayar, böylece uzaydaki yönelimi ön düzleme neredeyse dik olur. Bu nedenle, 0,02 s vektörünün standart uçların ekseni üzerindeki izdüşümü sıfıra yaklaşır ve dişlerin kendileri Q kayıtlı değil.
Son moment vektörü (0,06 s) daha da yukarı doğru kayar ve üç standart uzuv ucunun hepsinin eksenlerinin negatif kısımlarına yansıtılmaya başlar, bu da oldukça derin dişlerin ortaya çıkmasına neden olur S v S u Ve SM.
Bu nedenle kalbin enine eksen etrafındaki dönüşünü belirlemek için kompleksin konfigürasyonunun değerlendirilmesi gerekir. QRS standart uzuv kablolarında.
Atriyal Dalga Analizi R
Kalbin ön-arka, uzunlamasına ve enine eksenler etrafındaki dönüşünü belirledikten sonra atriyal dalganın analizine geçin. R. Uç analizi Rşunları içerir: 1) diş genliğinin ölçümü R, 2) diş süresi ölçümü RU 3) diş polaritesinin belirlenmesi R, 4) diş şeklinin belirlenmesi R.
Uç genliği R kontur çizgisinden sivri ucun tepesine kadar ölçülür ve süresi gösterildiği gibi sivri ucun başından sonuna kadardır

incirde. 4.24. Normal diş genliği R 2,5 mm'yi geçmez ve süresi 0,1 saniyedir. Uç polaritesi R I, II ve III'teki derivasyonlarda, uyarma dalgasının atriyum boyunca hareket yönünü ve dolayısıyla uyarma kaynağının (kalp pili) lokalizasyonunu gösteren en önemli elektrokardiyografik işarettir. Hatırlayacağınız gibi, uyarma dalgasının atriyum boyunca yukarıdan aşağıya ve sola doğru normal hareketi ile dişler pozitiftir ve uyarım aşağıdan yukarıya doğru yönlendirildiğinde negatiftir.Bu ikinci durumda, kalp pili atriyumun alt kısımlarında veya AV. düğümün üst kısmında bulunur. Sağ atriyumun orta kısmından çıkan uyarı ile depolarizasyon dalgası hem yukarı hem de aşağı doğru yönlendirilir. Ortalama vektör R sırasıyla sola doğru yönlendirilmiş diş R ( arttıkça diş büyür Plv ve P dalgası ||(negatif ve sığ hale gelir.
Diş şeklinin belirlenmesi büyük pratik öneme sahiptir R.İki apeks ile bölünmüş, genişletilmiş diş R sol derivasyonlarda (I, aVL, V 5, V 6) mitral kalp defekti ve sol atriyal hipertrofisi olan ve sivri yüksek amplitüdlü dişleri olan hastalar için tipiktir R I, III, aVF derivasyonlarında kor pulmonale hastalarında sağ atriyumun hipertrofisi gözleniyor (daha fazla ayrıntı için Bölüm 7'ye bakın).
Ventriküler kompleks analizi QRST
Çiçeklenmeçiçek taşıyan bir sürgün veya sürgün sistemidir. Çiçeklenme eksenlerinin düğümlerinde, sürgünün bitkisel kısmındakiyle aynı yapraklar veya fotosentez yapma yeteneğini kaybetmiş değiştirilmiş yapraklar vardır - diş telleri ve pediselin düğümlerinde - diş telleri.
Çiçek salkımlarının tek çiçeklere göre biyolojik avantajı, tozlaşma garantisini arttırmak ve yavaş yavaş çiçeklenmeleri nedeniyle olumsuz çevresel faktörlerin çiçeklere zarar verme olasılığını azaltmaktır. Bitkilerin çoğunda çiçek salkımına sahiptir.İki tür çiçeklenme vardır: karmaşıkçiçekler ana eksenin dallarında bulunduğunda ve basit, pedicelli veya pedicelsiz çiçekler doğrudan ana eksene yerleştirildiğinde.
Eksenlerin büyüme yöntemine bağlı olarak karmaşık salkımlar ikiye ayrılır: sempozyum(tanımlanmış) - eksen bir çiçekle biter, çiçekler üstten yan dallara doğru veya çiçekler aynı düzlemde bulunuyorsa merkezkaç şeklinde çiçek açar; Ve tek ayaklı(belirsiz) - eksen süresiz olarak büyür, çiçekler tabandan üste doğru veya çiçekler aynı düzlemde bulunuyorsa merkezcil olarak çiçek açar.
Karmaşık sempozyum salkımları:
- monochasium - ana eksen bir çiçekle biter; altında ikinci dereceden bir eksen oluşturulur, yine bir çiçek vb. ile taçlandırılmıştır; çiçek altı eksenleri bir yönde hareket ederse, o zaman bir kıvrılma oluşur, ancak dönüşümlü olarak bir yönde veya diğerinde ise bir girus oluşur; yan dalların kısaltıldığı kıvrıma glomerulus denir;
- dikazya - çiçeğin altında, her biri aynı zamanda bir çiçekle biten ve aynı dallanma yöntemini tekrarlayan iki alt çiçek ekseni üreten, vb. iki zıt eksen oluşturulur;
- pleiochasia - ana eksenden ikiden fazla alt çiçek ekseni uzanır, bir apikal çiçek taşır ve bir monochasia veya dikazi sarmalı oluşturur;
- thyrsus - yukarıdaki çiçek salkımları ana eksende bulunur, çoğunlukla piramidal bir şekle sahiptir.
Karmaşık monopodial çiçek salkımları:
- salkım - çok dallı bir çiçeklenme, alt yan dallar üst dallardan daha kuvvetli dallanır;
- corymb - çiçeklerin aşağı yukarı aynı düzlemde bulunduğu bir salkım;
- karmaşık başak - ana eksen, üzerinde pedicel olmayan çiçeklerin bulunduğu dalları oluşturur; bu dallara spikeletler denir;
- karmaşık şemsiye - ikinci dereceden eksenler arasındaki mesafeler kısalır ve birinci dereceden eksenin tepesinden uzaklaşır; pediceller arasındaki mesafeler kısalır ve ikinci derece eksenin üst kısmında bulunurlar; Çoğu zaman, ikinci dereceden eksenlerin tabanındaki yapraklar genel bir kıvrım oluşturur ve pedisellerin tabanında özel bir kıvrım oluşturur.
Listelenenlere ek olarak, adı verilen karmaşık çiçek salkımları da vardır. agrega. Farklı çiçek salkımlarının birleşiminden oluşurlar. Örneğin, civanperçemi toplu bir çiçeklenmeye sahiptir - bir kalkan içinde toplanan sepetler; fescue ve bluegrass'ta bir salkım halinde toplanmış başakçıklar bulunur.
Uzun bir eksene sahip basit salkımlar:
- salkım - ana eksen, genellikle aynı uzunlukta pedicelli çiçekler taşır;
- başak - ana eksen pedicelsiz çiçekler taşır;
- kedicik - sarkık bir kulak, yani yumuşak eksenli bir kulak; çiçeklenmeden sonra çiçeklenme genellikle düşer;
- koçanı - çok kalınlaştırılmış bir eksene sahip, bir veya daha fazla yaprakla çevrelenmiş, sözde spathe veya kanat olan bir kulak.
Kısaltılmış eksenli basit salkımlar:
- şemsiye - ana eksenin kısaltıldığı ve neredeyse aynı uzunluğa sahip olan pedicellerin eksenin üstünden uzandığı bir çiçeklenme;
- kafa - çiçekleri pedicelsiz veya çok kısa olan bir şemsiye;
- sepet - ana eksenin üst kısmı, üzerinde sıkıca kapatılmış çiçeklerin bulunduğu bir yatak şeklinde büyür; apikal yapraklar kalabalıktır ve bir kıvrım oluşturur.
Cymoid salkımına büyük bir çiçek salkımına sahip gruptur. Cymoid salkımları arasında iki ana tip vardır: simoidler ve tirsiler. Simoidler genellikle basitleştirilmiş tirsilerdir. Thyrsae dallanmış çiçek salkımlarıdır ve dallanma derecesi tabandan tepeye doğru azalır. Tirsusun ana ekseni tek ayaklı olarak büyür, ancak bir veya başka bir düzenin kısmi salkımları simoidlerdir.
Ana ekseni çiçekle biten Thyrsa'lara kapalı denir, aksi takdirde açık kabul edilir. Yan eksenlerin dallanma derecesine bağlı olarak, simoidlerin üçüncü ve daha yüksek derecelerin eksenlerinde yer aldığı pleiothyrsus ayırt edilir; simoidlerin ikinci derecenin eksenlerinde yer aldığı ditirler; ve simoidlerin doğrudan çiçeklenmenin ana ekseni üzerinde yer aldığı monothyrsus.Tyrsi'nin rasem, başak, kedicik, şemsiye veya başlık ile dışsal benzerliği salkım şeklinde, başak şeklinde, kedicik şeklinde konuşmamızı sağlar, şemsiye şeklinde, kapitat thyrsi vb. Thyrsae, yan eksenlerin sayısını azaltarak (azaltarak), internodları kısaltarak vb. Kolayca diğer çiçek salkımına dönüştürülür. Thyrsae bitkilerde çok sık bulunur. Örneğin thyrsus at kestanesinin çiçeğidir. Çeşitli türlerdeki Thyrs'ler birçok Lamiaceae'nin çiçek salkımını temsil eder. Huş ağacı salkımları kedicik şeklindeki tirsuslardır.
Saz alt ailesi (Caricoideae), üç kahverengi puldan oluşan Sibirya kobresisi (Kobresia sibirica) hariç, periantı olmayan tek cinsiyetli çiçeklerle karakterize edilir (Şekil 167). Bu ailenin temsilcilerinin başakçıkları, pullu örtücü yaprakların dingillerinde bulunur ve başakçının ventral (ventral) tarafında bulunan ve bitkisel sürgünün ön yaprağına homolog olan pullarla çevrilidir. Schoenoxiphium ve Kobresia cinslerinde bu pulların kenarları kaynaşmamış veya yarısına kadar (nadiren tepeye kadar) kaynaşmış kenarlara sahiptir ve Uncinia ve Carex cinslerinde ise oldukça başkalaşmış, kapalı ve kese şeklindedirler. Schenoxyphium ve cobresia genellikle uzun (birinci cinste) ve çok kısaltılmış, zar zor gelişmiş (ikinci cinste) eksene sahip biseksüel spikeletlere sahiptir. Uncinia ve sazda başakçıklar, kese adı verilen kese benzeri bir pulun içine alınmış tek bir dişi çiçeğe indirgenir. Uncinia türlerinde dişi çiçek, hâlâ korunmuş olan başakçıkların tabanında bulunur; ikincisi sazların büyük çoğunluğunda tamamen azaltılmıştır. Stamenler 3. Stil 2-3 stigmatik dallıdır. Alt aile, 4 cins içeren bir saz kabilesinden (Cariceae) oluşur. Bunlardan en ilkel olanı, 2'si Madagaskar'da ve geri kalanı Güney Afrika'da yetişen yaklaşık 15 türden oluşan Schenoxyphium'dur. Dağlarda, nemli ve bataklık yerlerde yetişirler. Bunlar bazal, genellikle düz yapraklı ve başak şeklinde veya panik şeklinde çiçek salkımına sahip çok yıllık bitkilerdir. Başakçıklar değişen derecelerde kapalı, bazen kese benzeri pullarla çevrilidir. İçinde, tabanında dişi bir çiçeğin oturduğu, spikeletin kuvvetle düzleştirilmiş bir ekseni vardır ve üst kısmında birkaç erkek bulunur (Şek. 167). Bazı türlerde eksende erkek çiçekler bulunmaz; ikincisi aynı çiçeklenme döneminde erkek spikelets oluşturur.
Saz alt ailesinin bir sonraki cinsi - Uncinia - esas olarak güney yarımkürede (Afrika hariç) dağıtılan 40-50 türden oluşur. Türlerin çoğu (32) Yeni Zelanda'ya özgüdür. Ekvatorun kuzeyinde, uncinia Güneydoğu Asya'da (Kalimantan Adası, Filipinler), Hawaii Adalarında, Orta ve Güney Amerika'da ve Batı Hint Adaları'nda bulunur. Bu cinsin türleri deniz seviyesinden alp kuşağına kadar, bataklık çayırlarda, bataklıklarda, nemli ormanlarda ve çalılıklarda sıklıkla çalılık oluşturdukları yerlerde yetişir. Uncinia türleri çok yıllık, genellikle bazal doğrusal yaprakları olan yoğun çimenli bitkilerdir. Çiçeklenme, üst kısımda erkek çiçekleri taşıyan apikal bir başaktır ve alt kısımda dişi çiçekleri çevreleyen keselerden oluşur ve buradan tepe noktasında kavisli kanca şeklinde bir başakçık ekseni çıkıntı yapar. Eksenin kanca şeklindeki tepe noktası, anatomik çalışmaların gösterdiği gibi sinüste az gelişmiş bir böbrek bulunan sert, sarılmış bir kaplama ölçeğidir. Stamenler 3. Stigma şeklinde 3 dallı stil. Meyve üçgen şeklindedir ve bir kese içine alınır. Keseler kanca şeklindeki eksenleriyle hayvanların kürklerine veya tüylerine tutunarak yayılırlar.
Cobresia cinsi, esas olarak kuzey yarımkürenin ılıman ve kısmen subtropikal ve tropik bölgelerinin dağlık bölgelerinde, ancak esas olarak Asya'da dağıtılan yaklaşık 40 tür içerir; az sayıda tür Kuzey Kutbu'na nüfuz eder. Cobresias, az yağış alan yaylaların karakteristik bitkileridir. Nehir ve göl kıyılarında, nemli ve bataklıklarda, genellikle tuzlu çayırlarda, çayır bozkırlarında ve kayalık yamaçlarda yetişirler. Bu cinsin türleri genellikle kobresia çayırlarını (cobresia çayırları veya fundalıklar olarak adlandırılır) oluşturur. Cobresias, düz veya fırça benzeri plakalara sahip bazal yaprakları olan çok yıllık yoğun çim otlarıdır. Çiçeklenme başak şeklinde veya daha az sıklıkla panikülattır, en üst kısmı bazen erkek olan küçük 2-7 çiçekli biseksüel spikeletlerden oluşur. Başakçığı çevreleyen pullar genellikle kaynaşmamış kenarlara veya nadiren yarıya kadar veya neredeyse tepeye kadar kaynaşmış kenarlara sahiptir. Spikeletler (eksenleri büyük ölçüde kısaltılmıştır, neredeyse ifade edilmez) bir dişi çiçek ve onun dışına doğru yerleştirilmiş 1-6 erkek çiçekten oluşur. İkincisi, kaplama pullarının koltuğunda oturan 3 organdan oluşur. Stilus 3, nadiren 2 stigmatik dallıdır. Meyve genellikle üçgen şeklindedir. Cobresias tohumlarla çoğalır ve bunlardan yalnızca biri - güçlü kobresia (K.robusta) - aynı zamanda vejetatif olarak, sürünen rizomlar yoluyla. Meyveler otçul memeliler tarafından dağıtılır. Tien Shan ve Pamir-Alai'de geniş alanları kaplayan Cobresia tüylü (K. capilliformis) ve diğer bazı türler değerli mera bitkileridir.

Saz cinsi (Carex) yaklaşık 1500 (diğer kaynaklara göre 2000 - 2500'e kadar) türe sahiptir ve nispeten az sayıdaki en büyük anjiyosperm cinsine aittir. Saz türleri dünya genelinde yaygındır, ancak esas olarak ılıman ve soğuk bölgelerde, yalnızca nemli ve bataklık değil, aynı zamanda kuru habitatların bitki örtüsünün bileşiminde de önemli bir rol oynarlar. Yaklaşık 400 tür sazın bulunduğu Sovyetler Birliği'nin bitki örtüsüne katılım derecesi açısından, çimenler ve asteraceae'den sonra ikinci sırada yer alıyorlar. Bu cinsin pek çok türü, bozkır ve hatta yarı çöl ve çöl bitkileri de dahil olmak üzere çeşitli bitki topluluklarının düzenleyicileridir. Sazlar deniz seviyesinden yüksek dağlara kadar hemen hemen her yerde yetişir. Türlerin çoğu bataklıklarda, bataklık ve nemli çayırlarda ve su kütlelerinin kıyılarında yaşar (örneğin, şişe saz - C. rostrata, mesane saz - C. vesicaria, tilki saz - C. vulpina, Şekil 174, ikievli saz - C. dioica, Şekil 174). Diğer sazlar tropik yağmur ormanları da dahil olmak üzere farklı orman türlerinde bulunur. Bazı türler kuru çayırlarda, bozkırlarda ve kuru yamaçlarda yetişir (alçak saz - C. hnmilis, erken saz - C. paecox, bodur saz - C. supina vb.). Kalın sütunlu saz (C. pachystylis) killi yarı çöllerin karakteristik bir bitkisidir ve şişmiş saz (C. physodes) kumlu çöllerin karakteristik bir bitkisidir. İkinci tür, meyvelerle şişmiş, kuvvetli kabarcık benzeri, kırmızımsı paslı, çok büyük (2 cm uzunluğa kadar) keseler nedeniyle alışılmadık derecede orijinal bir görünüme sahiptir (Şek. 174). Deniz kıyılarında, kumullarda, kıyı çayırlarında, çakıl taşlarında ve kayalıklarda pek çok tür yaşar. Ilıman bölgedeki türlerin çoğu ovalarda yetişir, ancak bazı sazlar subalpin ve alpin çayırların tipik sakinleridir. Tropikal bölgelerde, özellikle de 66 saz türünün yetiştiği Malezya floristik bölgesinde, sazlar esas olarak dağlarda yaşar ve deniz seviyesinden deniz seviyesinden 500 m yüksekliğe kadar yalnızca 10 kadar tür yetişir.
Sazlar çok yıllık tek evcikli, çok nadiren iki evcikli, sürünen veya kısaltılmış rizomlara sahip, tutamlar veya bazen tümsekler oluşturan bitkilerdir. Yaprakların tamamı genellikle bazaldir, çok daha az sıklıkla, neredeyse tüm gövde boyunca yer alan düğümlerden uzanan kök yaprakları da vardır; Yaprak ayası genellikle doğrusaldır, nadiren mızrak şeklindedir (örneğin, paslı benekli sazda - C. siderosticta, Şekil 175) ve bazen bir yaprak sapı şeklinde daralmıştır. Çiçekler tek eşeylidir; başak şeklinde, paniküler, salkım veya kapitat çiçeklenme oluşturan başakçıklar halindedir; bazen çiçeklenme tek başak şeklindedir. Başakçıklar tamamen erkek veya dişi çiçeklerden oluşur veya iki türden karışıktır: çift cinsiyetli - başakçıkların üst kısmında erkek çiçekler ve alt kısmında dişi çiçekler bulunur; gynecandrous - dişi çiçekler üstte bulunur ve erkek çiçekler alt kısımda bulunur. alt. Dişi çiçekler, meyveyle birlikte artan, tepe noktasında daralan ve stilin bir kısmının ve stigmatik dalların çıktığı bir ağızlığa dönüşen bir kese içine alınır. Yukarıda, keseler içine alınmış sazların dişi çiçeklerinin azaltılmış, tek çiçekli spikeletler olduğu söylenmişti. Bu nedenle, aynı eksen üzerinde oturan keselerden oluşan bir koleksiyon karmaşık bir sivri uçtur. Erkek çiçek koleksiyonları basit sivri uçlardır. Bununla birlikte, kolaylık sağlamak için, sazların hem karmaşık hem de basit sivri uçlarına genellikle basit başakçıklar denir. Keselerin karakterleri son derece çeşitlidir ve sazların taksonomisi açısından büyük önem taşır. Bu cinsin çok sayıda türü esas olarak keselerinde farklılık gösterir. Keseler şüphesiz önemli biyolojik öneme sahip bir organdır. Belki de saz cinsinin ailenin tüm cinsleri arasında en çok sayıda ve en yaygın olanı haline gelmesi kesenin ortaya çıkması sayesinde olmuştur. Keseler, yumurtalığı ve gelişen meyveyi olumsuz koşullara maruz kalmaktan korur ve birçok türde aynı zamanda meyvelerin dağılımına da bir adaptasyon sağlar. Sazların stilinde genellikle 2 - 3 uzun stigmatik dal bulunur. Meyveler üçgen veya hafif bikonvekstir.
Birçok saz ilkbaharda erken çiçek açar. Bu cinsin türleri anemofil bitkilerdir, ancak bazılarının görünüşe göre entomofili de sergilediği görülmektedir. Bu nedenle, en erken çiçek açan türlerden ikisi olan funda sazının (C. ericetorum) ve karanfil sazının (C. caryophyllea) polen toplamak için bal arıları tarafından düzenli olarak ziyaret edildiği ve görünüşe göre aynı zamanda çapraz tozlaşma da gerçekleştirdiği bilinmektedir. Saz çiçeklenmesinin biyolojisi hakkında neredeyse hiçbir bilgi yoktur. Çiçekleri protogin olarak kabul edilir. Ancak tüylü sazın (C. pilosa) çiçeklenmesine ilişkin özel gözlemlerin gösterdiği gibi, morfolojik protoji, fizyolojik ile örtüşmeyebilir (L. Antonova, 1976). Bu türde, tam gelişmiş stigmaların ortaya çıkışı, anterlerin açılmasından önce gerçekleşir. Bununla birlikte, stigma fizyolojik olgunluğa, yani poleni algılama yeteneğine ancak anterler açıldığında ve polen dağıldığında ulaşır. Bu nedenle tüylü saz aslında prototip değil, homojen bir bitkidir. Kendi kendine tozlaşma muhtemelen kısmi kendi kendine uyumsuzluk nedeniyle hariç tutulmuştur. Kendi kendine tozlaşma ile bu türün çapraz tozlaşmaya göre önemli ölçüde daha az meyve ürettiği deneysel olarak tespit edilmiştir. Sazlar tohumlarla çoğalır, ancak sürünen rizomlu türlerde bitkisel çoğalma belirgin şekilde baskındır. Meyvelerin dağıtımı çeşitli şekillerde gerçekleşir. Tabanda şişmiş veya süngerimsi keseler içinde bulunan kıyı sazlarının meyveleri su yoluyla dağıtılır. Şişmiş saz keseleri anemokoriye iyi adapte olmuştur. Hafif rüzgarlarda bile kum yüzeyinde kolaylıkla yuvarlanırlar. Pek çok türde keseler, çoğunlukla kuşların ayaklarındaki kirle birlikte, epizoochorous olarak dağıtılır. Bazı sazların keseleri kuşların tüylerine de yapışabilir. Ördekler tarafından yutulan meyvelerin endozoochor dağılımı önemli bir rol oynar. Kuşlar, tropikal bir tür olan saz meyvesinin turuncu-kırmızı, biraz etli keselerini taşırlar (C. baccans, Şekil 175). Palmiye sazlığı (C.digitata) ve ornithopoda sazlığı (C. ornithopoda) gibi bazı orman sazlıklarının uzun etli tabanlara sahip keseleri karıncalar tarafından taşınır.
Kalp döndüğünde Apeks enine ekseni etrafında öne doğru ilerlerken, ortalama QRS vektörü ileri doğru sapar, başlangıç vektörü (Q) normalden daha fazla sağa ve yukarıya doğru yönlendirilir (F düzleminde). Ön düzleme paralel olarak yerleştirilmiştir ve bu nedenle tüm standart kabloların (I, II ve III) eksi eksenlerine açıkça çıkıntı yapar.
Açık EKG belirgin bir dalga QI, II, III kaydedilir. Son vektör (S), frontal düzleme dik olarak geriye ve aşağıya doğru sapar ve standart derivasyonların ekseninde eksiye projeksiyon yapmaz, bu nedenle S dalgası I, II, III derivasyonlarında kaydedilmez. EKG'de I, II ve III'teki enine eksen etrafında apeksi öne doğru dönerek qR kompleksini kaydeder.
Kalp döndüğünde Tepe noktası enine eksen etrafındayken, ortalama QRS vektörü geriye doğru (S düzleminde) sapar, son vektör (S) sağa ve yukarıya doğru saparak I. uçların eksenlerinin negatif kutbuna önemli bir projeksiyon verir, II ve III. EKG'de belirgin bir SI, II, III dalgası görülüyor. Başlangıç vektörü (Q) aşağıya ve ileriye doğru yönlendirilir ve bu nedenle standart uçların eksenlerinin negatif kutbuna yansıtılmaz. Sonuç olarak EKG'de I, II ve III'te Q dalgası yok, QRSI, II, III kompleksi RS tipi ile temsil ediliyor.
EKG sağlıklı kadın D., 30 yaşında. Sinüs ritmi düzenli olup dakikada 67'dir. P - Q=0,12 sn. P = 0,10 sn. QRS = 0,08 sn. Q - T = 0,38 sn. Ru>RI>Rir AQRS=+52°. Ap=+35°. =+38°'de. Kompleks QRSI,II,III tip qR. Bu, başlangıç vektörünün (Q) sağa ve yukarıya normalden daha fazla yönlendirildiğini ve bu nedenle tüm standart uçların (dalga qI, II, III) eksisine yansıtıldığını gösterir. Son vektör (S), ön düzleme dik olarak geriye ve aşağıya doğru sapar ve I, II, III derivasyonlarının eksenlerine yansıtılmaz (S dalgası yoktur, cw). Başlangıç ve son vektörlerin yönündeki bu tür değişiklikler, kalbin tepe noktası öne doğru dönmesinden kaynaklanıyor olabilir. QRS geçiş bölgesinin normal konumunun sağ sınırı olan derivasyon V2 ile çakıştığı unutulmamalıdır. Uzunlamasına eksen etrafında saat yönünde eşzamanlı hafif bir dönüşü yansıtan karmaşık QRSV5V6 tip RS. Tüm derivasyonlarda P, T dalgaları ve RS-T segmenti normal.
Çözüm. Normal bir EKG'nin bir çeşidi (kalbin tepe noktası enine eksen etrafında ileri ve uzunlamasına eksen etrafında saat yönünde olacak şekilde dönmesi).
Sağlıklı bir adamın EKG'si K., 37 yıl. Şiddetli sinüs bradikardisi, 1 dakikada 50. Aralık P - Q=0,15 sn. P = 0,11 sn. =0,09 sn. Q - T=0,39 sn. RII>RI>RIII. AQRS = +50°. Ar=+65°. =+50°'de. QRS açısı - T=0°. Kompleks QRSI,II,III tip qR. Q dalgası en çok derivasyon II'de belirgindir; amplitüdü 3 mm'dir ve süresi 0,03 saniyeden biraz daha azdır. (normal boyutlar). Tanımlanan QRS şekli, kalbin tepe noktasını öne doğru döndürmesiyle ilişkilidir.
Göğsünde potansiyel müşteriler QRSV5, V6 kompleksi de qR tipindedir ve RV1 dalgası belirgindir ancak genişlememiştir (genlik 5 mm). Bu QRS değişiklikleri, kalbin uzunlamasına ekseni etrafında saat yönünün tersine döndüğünü gösterir. Geçiş bölgesi normal olarak bulunur (V2 ve V3 arasında). Geriye kalan EKG dalgaları normaldir. RS segmenti - TII,III 0,5 mm'den fazla yükselmez, bu normal olabilir.
Çözüm. Sinüs bradikardisi. Kalbin saat yönünün tersine ve tepe noktası öne doğru döndürülmesi (normal EKG'nin bir çeşidi).
Sağlıklı bir kadının EKG'si K., 31 yaşında. Sinüs ritmi düzenli olup dakikada 67'dir. P - Q=0,16 sn. P=0,09 sn. QRS=0,08 sn. Q - T=0,39 sn. RII>RI>RIII. AQRS=+56°. =+26°'de. QRS açısı - T=30°. Ar=+35°.
Karmaşık QRSI,II,III tipi Rs. I, II, III'teki derivasyonlarda telaffuz edilen S, son vektörün (S) sağa ve yukarıya doğru önemli bir sapmasını gösterir. QI, II, III dalgasının yokluğu, başlangıç QRS vektörünün aşağı ve ileri yönü (standart derivasyonların pozitif kutbuna doğru) ile ilişkilidir. Başlangıç ve son QRS vektörlerinin bu yönelimi, kalbin apeksi enine ekseni etrafında geriye doğru dönecek şekilde dönmesine bağlı olabilir (tip SI, SII, SIII EKG). Geri kalan EKG dalgaları olağan normal özellikler dahilindedir: QRSV6 tipi qR'ler. V2 ve V3 arasındaki QRS geçiş bölgesi, RS segmenti - TV2 1 mm yukarı kaydırılır. Kalan derivasyonlarda RS-T izoelektrik hat seviyesindedir, TIII hafif negatiftir, TaVF pozitiftir, TV1 negatiftir, TVJ_V6 pozitiftir ve V2V3'te biraz daha büyük genliğe sahiptir. P dalgası normal şekil ve büyüklüktedir.
Çözüm. Normal EKG tipi SI, SII, SIII'in bir çeşidi (kalbin tepe noktası enine eksen etrafında geriye doğru olacak şekilde dönmesi).

Daha az yaygın olarak, EKG, kalbin ön-arka (sagital) düzlemde meydana gelen enine ekseni etrafındaki rotasyonlarını kaydeder (Şekil 57). Kalbin enine eksen etrafındaki rotasyonları genellikle kalbin tepe noktasının normal konumuna göre ileri veya geri sapması ile ilişkilidir, bu da sagittaldeki ventriküler depolarizasyonun üç moment vektörlerinin olağan mekansal düzenlemesinin ihlaline yol açar. ve ön düzlemler.
Kalbin apeks ileri veya geri olacak şekilde transvers eksen etrafındaki rotasyonları en iyi üç standart ekstremite derivasyonunda kaydedilir. Şek. 57. Gözlemciye belirli bir açıyla döndürülmüş tanıdık altı eksenli Bailey koordinat sisteminin yanı sıra üç moment vektörünün (0,02 s, 0,04 ve 0,06 s) uzamsal düzenlemesini gösterir.
İÇİNDE
Pirinç. 57. Üç standart derivasyonda EKG formu. a - kalbin normal pozisyonunda, b - kalp, tepe noktası öne gelecek şekilde enine eksen etrafında döndürüldüğünde; c – kalp, apeks geriye doğru olacak şekilde enine eksen etrafında döndürüldüğünde.
Her iki vektör de uzaysal olarak ön düzleme belirli bir açıda yerleştirilmiştir; 0,02 s'lik vektör ileri doğru yönlendirilir ve 0,06 s'lik vektör geriye doğru yönlendirilir. Her iki vektör de standart derivasyonların eksenlerinin negatif kısımlarına yansıtılır, bunun sonucunda bu derivasyonlarda nispeten küçük genlikli Q ve S dalgaları kaydedilebilir.Q ve S dalgalarının yalnızca bir tanesine kaydedilebileceği unutulmamalıdır. veya üç standart derivasyondan ikisi: I ve II veya II ve III'te.
Kalp, apeks ileri doğru olacak şekilde enine eksen etrafında döndüğünde (Şekil 57, b), başlangıçtaki tork vektörü (0,02 s) daha da yukarıya ve bir miktar sağa doğru kayar ve bu nedenle Q dalgası her üçünde de kaydedilmeye başlar. standart öne çıkıyor ve daha belirgin hale geliyor.
Son moment vektörü (0,06 s) aşağı ve geriye doğru sapar, bunun sonucunda artık ön düzleme neredeyse dik olarak konumlandırılır. Bu nedenle tüm standart derivasyonların eksenlerindeki projeksiyonu sıfıra yaklaşır ve bu da S dalgasının bu derivasyonlarda kaybolmasına yol açar.
HATIRLAMAK! Kalp apeks öne gelecek şekilde transvers eksen etrafında döndüğünde, standart derivasyonlardaki ventriküler QRS kompleksi qR I, qR II ve qR III formunu alır.
Kalp, tepe noktası geriye doğru olacak şekilde enine eksen etrafında döndüğünde (Şekil 57, c), başlangıç moment vektörü (0,02 s), uzaydaki yönelimi ön düzleme neredeyse dik olacak şekilde ileri ve aşağı kayar. Bu nedenle, 0,02 s vektörünün standart uçların ekseni üzerindeki izdüşümü sıfıra yaklaşır ve Q dalgalarının kendileri kaydedilmez.
Son moment vektörü (0,06 s) daha da yukarı doğru kayar ve üç standart uzuv ucunun hepsinin eksenlerinin negatif kısımlarına yansıtılmaya başlar, bu da oldukça derin S I, S II ve S III dişlerinin ortaya çıkmasına yol açar.
HATIRLAMAK! Kalp, apeks geriye doğru enine eksen etrafında döndüğünde, standart derivasyonlardaki ventriküler kompleks RS I, RS II, RS III formunu alır.
Bu nedenle kalbin transvers eksen etrafındaki rotasyonunu belirlemek için standart ekstremite derivasyonlarındaki QRS kompleksinin konfigürasyonunu değerlendirmek gerekir.
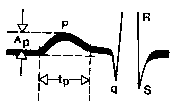
Pirinç. 58. EKG'nin P dalgasının genliğinin ve süresinin ölçülmesi. A p – P dalgasının genliği, t p – P dalgasının süresi.